シナプスの歴史
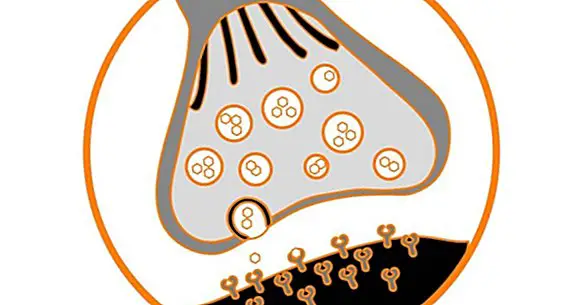
脳は、シナプスとして知られている小さな空間によって分離されているそのニューロン間の何千との相互接続を含む。 これは、情報の伝達がニューロンからニューロンへと進むところである .
しばらくの間、シナプスの活動は静的ではなく、常に同じではないことが分かっています。それは、私たちが生きるもののような外部刺激の結果として増強されるか、または減少され得る。シナプスを調節することができるこの品質は、大脳可塑性または神経可塑性として知られている。
今まで、シナプスを調節するこの能力は学習と記憶のような脳の発達にとって重要な2つの活動に積極的に関与していると考えられてきた。私はこれまで説明してきました。 記憶の機能を理解するためにシナプスはあまり重要ではない それは通常信じられている通りです。
シナプスの歴史
Ramon Y Cajalのおかげで、我々はニューロンが統一された組織を形成しないことを知っていますが、それらのすべては、後にSherringtonが "シナプス"と呼ぶ顕微鏡の場所である、数十年後、心理学者のDonald Hebbは、シナプスが時間的には必ずしも同じではなく、変調することができる理論、すなわち、我々が神経可塑性として知っていることを語った 2つ以上のニューロンは、それらの間の関係を強化または劣化させる可能性がある 特定の通信チャネルを他の通信チャネルより頻繁にする。この理論を適用する50年前の興味深い事実として、Ramóny Cajalは彼の著書にこのモジュレーションの存在の証拠を残しました。
今日我々は、脳の可塑性の過程で使用される2つのメカニズム、すなわち、2つのニューロン間のシナプスの強化である長期増強(LTP) (LTD)、これは情報の伝達の減少である第1のものとは反対である。
記憶と神経科学、論争を伴う経験的証拠
学習は、新しい知識を得るために、私たちが人生における物事や出来事を関連づけるプロセスです。記憶は、時間の経過とともに学習されたこの知識を維持し、保持する活動である。歴史を通して、何百もの実験が、脳がこれらの2つの活動をどのようにして行うかを調べるために行われてきた。
この研究の古典は、小さな無脊椎動物(Aplysiaとして知られる海洋のカタツムリ)を持つKandel and Siegelbaum(2013)の研究です。この調査では、 彼らは、動物が環境にどのように反応するかの結果としてシナプス導電率の変化が生じることを見出した シナプスが学習と記憶の過程に関与していることを示しています。しかし、ChenらによるAplysiaを用いた最近の実験では、 (2014年)は、以前に到達した結論と衝突する何かを発見した。この研究は、シナプスが薬物によって阻害された後、運動機能の動物において長期記憶が持続することを示し、シナプスが全記憶過程に関与するという考えに疑問を投げかけている。
この考えを支持する別のケースは、Johanssonらによって提案された実験から生じる。 (2014)。この機会に、小脳のプルキンエ細胞を研究した。これらの細胞は、運動のリズムを制御し、直接的に刺激され、薬剤によるシナプスの抑制下で、すべての予後に対して、そのペースを設定し続けた。 Johanssonは、彼の記憶が外部のメカニズムの影響を受けていないこと、そしてシナプスの影響とは無関係にメカニズムを個別に制御するのはプルキンエ細胞そのものであると結論づけた。
最後に、Ryanらによるプロジェクト(2015)はシナプスの強さが記憶の統合における重要なポイントではないことを実証しました。彼の研究によると、動物に蛋白質阻害剤を注射すると、逆行性健忘症が生じる、すなわち、彼らは新しい知識を保持することができない。しかし、この同じ状況では、特定のタンパク質(オプトジェネティックスとして知られている方法)の産生を刺激する光の小さな点滅を適用すると、誘発された化学的遮断にもかかわらず記憶を保持することができます。
学習と記憶、統一された、あるいは独立したメカニズム?
何かを覚えるには、まずそれについて学ぶ必要があります 。私はそれが原因であるかどうかわかりませんが、現在の神経科学文献はこれら2つの用語をまとめる傾向があり、それらが基礎としている実験は通常、学習過程と記憶を区別できない曖昧な結論をもち、共通のメカニズムであるか否か。
良い例は、学習センターとしての海馬の研究におけるMartin and Morris(2002)の研究である。この研究拠点は、神経伝達物質のグルタミン酸を認識し、LTPシグナルに関与するタンパク質であるN-メチル-D-アスパラギン酸(NMDA)の受容体に焦点を当てた。彼らは、視床下部の細胞に長期間の増強がなければ、新しい知識を学ぶことは不可能であることを実証した。実験は、阻害剤のないラットとは異なり、ラフトにラットを入れたラットにNMDA受容体ブロッカーを投与し、ラフトを入れてラフトの位置を知ることができないラフトを投与することからなっていた。
その後の研究は、ラットがインヒビターの投与前に訓練を受けた場合、ラットはLTPの喪失を「補償」する、すなわち記憶を有することを明らかにする。我々が示したいと思う結論は、 LTPは学習に積極的に参加していますが、情報検索ではそうはっきりしません .
脳の可塑性の意味
それを示す多くの実験があります 新しい知識の獲得に積極的に参加する神経可塑性 例えば上記の場合、または動物の学習を著しく妨げるグルタミン酸産生遺伝子が排除されたトランスジェニックマウスの作製において使用することができる。
その代わりに、あなたの記憶の中でのあなたの役割は疑う余地があります。記憶のメカニズムは、シナプスではなく細胞の内部にあるという理論が浮上し始めています。しかし、心理学者で神経科学者ラルフ・アドルフが指摘しているように、 神経科学は、学習と記憶が今後50年間にどのように働くかを解決する つまり、時間だけがすべてを明確にします。
書誌事項:
- Chen、S.、Cai、D.、Pearce、K.、Sun、P.Y.-W.、Roberts、A.C.、Glanzman、D.L。(2014)。 Aplysiaにおける行動およびシナプスの発現の消去後の長期記憶の回復。 eLife 3:e03896。 doi:10.7554 / eLife.03896。
- Johansson、F.、Jirenhed、D.A.、Rasmussen、A.、Zucca、R.およびHesslow、G.(2014)。小脳プルキンエ細胞に局在する記憶トレースおよびタイミング機構。 Proc。 Natl。 Acad。 Sci。U.S.A. 111,14930-14934。 doi:10.1073 / pnas.1415371111。
- Kandel、E.R.、およびSiegelbaum、S.A。(2013)。 ERニューロサイエンス、第5版、エド・カーン、JHシュワルツ、ジェイセル、シーゲルバムSA、およびAJ・ハドスペス(New York、NY:McGraw-Hill)の「暗黙の記憶記憶の細胞機構および個体の生物学的基礎」 )、1461-1486。
- Martin、S.J.およびMorris、R.G.M。(2002)。古いアイデアの新しい人生:シナプス可塑性と記憶仮説が再考された。 Hippocampus 12,609-636。 doi:10.1002 / hipo.10107。
- Ryan、T.J.、Roy、D.S。、Pignatelli、M.、Arons、A.、and Tonegawa、S.(2015)。エングラム細胞は、逆行性記憶失調症の記憶を保持する。 Science 348、1007-1013。 doi:10.1126 / science.aaa5542。